
不论是单细胞生物或组成多细胞有机体的每一个细胞,在它们的生命过程中,都会不断受到来自外部环境的各种理化因素的影响。在多细胞动物,由于绝大多数细胞是生活在直接浸浴它们的细胞外液、即内环境之中,因此出现在内环境中的各种化学分子,是它们最常能感受到的外来刺激:这不仅是指存在于细胞外液中的激素或其他体液性调节因子;而且就是在神经调节过程中,当神经信息由一个神经元向其他神经元传递或由神经元传给它的效应器细胞时,在绝大多数情况下,也都要通过一种或多种神经递质和调质为中介,通过这些化学分子在距离极小的突触间隙液中的扩散,才能作用到下一级神经元或效应器细胞。尽管激素和递质(或调质)等分子作为化学信号在细胞外液中播散的距离和范围有所不同,但对接受它们影响的靶细胞并不存在本质的差别。
细胞外液中的各种化学分子,并不需要自身进入它们的靶细胞后才能起作用(一些脂溶性的小分子类固醇激素和甲状腺激素例外,详见第十一章)它们大多数是选择性地同靶细胞膜上具有特异的受体性结构相结合,再通过跨膜信号传递(transembrane signaling)或跨膜信号转换(transmembranesognal transduction)过程,最后才间接地引起靶细胞膜的电变化或其他细胞内功能的改变。
机体和细胞也可能受到化学信号以外的其他性质的刺激,如机械的、电的和一定波长电磁波等来自外界环境的刺激的影响;但在动物进化的过程中,这些刺激信号大都由一些在结构和功能上高度分化了的特殊的感受器细胞来感受,引起相应的感受器细胞出现某种电反应。仔细分析各种感受器细胞接受它们所能感受的某种特异刺激信号的过程时发现(如耳蜗毛细胞接受声波振动和视网膜光感受细胞接受光刺激等),它们也涉及到外来刺激信号的跨膜传递,即刺激信号也要先作用于膜结构中的感受性结构,才能引起感受器细胞的电变化和随后的传入神经冲动。
不论是化学信号中的激素分子和递质(包括数十种可能起调质作用神经肽类物质)分子,以及非化学性的外界刺激信号,当它们作用于相应的靶细胞时,都是通过为数不多、作用形式也较为类似的途径来完成跨膜信号传递的;这些过程所涉及的膜蛋白质也为数不多,在生物合成上由几类特定基因家族所编码;正因为如此,由每个特定基因家族所表达生成的蛋白质分子,在肽链的氨基酸排列顺序上有较大的相同性(或同源性,homogeneity),功能上也较为类似。因此,关于跨膜信号传递的研究,早已超出了递质或激素作用机制的范畴,成为细胞生理学中一个有普遍意义的新篇章。试想,人体细胞都具有相同的遗传基因,因而一个感光细胞或一个普通体细胞,通过细胞膜上类似的蛋白质,以类似的方式接受它们所受到的外来刺激,可引起细胞本身功能的改变;而且各种不同的细胞通过少数几类膜蛋白质和几种作用方式,就能接受多种多样可能遇到的外界刺激信号的影响,显然符合“生物经济”的原则。
一、由具有特异感受结构的通道蛋白质完成的跨膜信号传递
(一)化学门控通道
对这种跨膜信号的传递方式的研究,最早是从对运动神经纤维末梢释放的乙酰胆碱(Ach)如何引起它所支配的骨骼肌细胞兴奋的研究开始的。早已知道,当神经冲动到达神经末梢处时,先是由末梢释放一定数量的Ach分子,后者再同肌细胞膜上称为终板(指有细胞膜上同神经末相对的那部分膜,其中所含膜蛋白与一般肌细胞膜不同)处的“受体”相结合,引起终板膜产生电变化,最后引起整个肌细胞的兴奋和收缩。由于神经-肌接头处的“受体”也可同烟碱相结合,因而过去在药理学分类中称它为N-型Ach受体。80年代后期,我国学者李镇源发现α-银环蛇毒同N-型受体有极高的特异性结合能力又有人发现一些电鱼的电器官中有密集的这种受体蛋白质分子存在;再依靠70年代以来蛋白质化学和分子生物学技术的迅速发展,目前不仅已将这种蛋白质分子提纯,而且基本上搞清了它的分子结构和它们在膜中的存在形式。原来它是由4种不同的亚单位组成的5聚体蛋白质(图2-7),总分子量约为290kd;每种亚单位都由一种mRNA编码,所生成的亚单位在膜结构中通过氢键等非共价键式的相互吸引,形成一个结构为α2βγδ的梅花状通道样结构(图2-7,A和B),而其中的两个α-亚单位正是同两分子ACH相结合的部位,这种结合可引起通道结构的开放,其几何大小足以使终板膜外高浓度的Na+内流,同时也能使膜内高浓度的K+外流结果是使原来存在两侧的静息电位近于消失,亦即使该处膜内外电位差接近于0值,这就是终板电位,于是完成了Ach这种化学信号的跨膜传递,因为肌细胞后来出现的兴奋和收缩都是以终板电位为起因的。
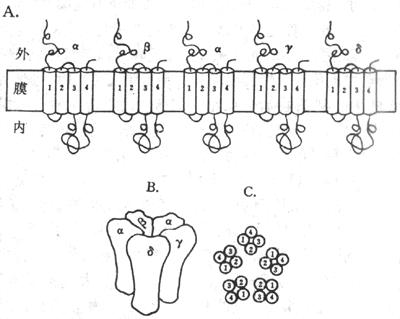
图2-7 N-型Ach门控通道的分子结构示意图
A:N-型Ach门控通的5个亚单位和它们所含α-螺旋在膜中存在形式的平面示意图
B:5个亚单位相互吸引,包绕成一个通道样结构 C:在跨膜通道结构中,各
个亚单位亿含α-螺旋在通道结构中的位置
用分子生物学实验技术证明,同其他膜结合蛋白质类似,在上述4种不同的亚单位肽链中,都存在有4种主要由20-25个疏水性氨基酸形成的α-螺旋,因而推测每个亚单位的肽链都要反复贯穿膜4次(图2-7,A),而5个亚单位又各以其第2个疏水性跨膜α-螺旋构成了水相孔道的“内壁”(图2-7C)。
由上述分子水平的研究成果可以知道,原初将终板膜上完成Ach跨膜信号传递的蛋白质称作“受体”是不符合实际情况的;它们是一种通道样结构,只是在组成通道的蛋白质亚单位中有两个亚单位具有同Ach分子特异地相结合的能力,并能因此引起通道蛋白质的变构作用而使通道开放,然后靠相应离子的易化扩散而完成跨膜信号传递。因此,这种蛋白质应称为N-型(或烟碱型)Ach门控通道,属于化学门控通道或化学依从性通道中的一种。
Ach在神经-肌接头处的跨膜信号传递机制的阐明,曾一度错误地推测,其他一些神经递质也都是以类似的方式作用于下一级神经元或相应的效应器细胞的;但后来的研究表明并非如此。目前只证明了一些氨基酸递质,包括谷氨酸、门冬氨酸、γ-氨基丁酸和甘氨酸等,主要是通过同N-型Ach门控通道结构类似的化学门控通道影响其靶细胞。
(二)电压门控通道
应用类似的技术,在80年代还陆续克隆出几种重要离子(如Na+、K+和Ca2+等离子)的电压门控通道,它们具有同化学门控能道类似的分子结构,但控制这类通道开放与否的因素,是这些通道所在膜两侧的跨膜电位的改变;也就是说,在这种通道的分子结构中,存在一些对跨膜电位的改变敏感的基团或亚单位,由后者诱发整个信道分子功能状态的改变。
在动物界,除了一些特殊的鱼类,一般没有专门感受外界电刺激或电场改变的器官或感受细胞,但在体内有很多细胞,如神经细胞和各种肌细胞,在它们的细胞膜中却具有多种电压门控通道蛋白质,它们可由于同一细胞相邻的膜两侧出现的电位改变而再现通道的开放,并由于随之出现的跨膜离子流而出现这些通道所在膜的特有的跨膜电位改变。例如,前述的终板膜由Ach门控通道开放而出现终板电位时,这个电位改变可使相邻的肌细胞膜中存在的电压门控式Na+通道和K+通道相继激活(即通道开放),出现肌细胞的所谓动作电位;当动作电位在神经纤维膜和肌细胞膜上传导时,也是由于一些电压门控通道被邻近已兴奋的膜的电变化所激活,结果使这些通道所在的膜也相继出现特有的电变化。由此可见,电压门控通道所起的功能,也是一种跨膜信号转换,只不过它们接受的外来刺激信号是电位变化,经过电压门控通道的开闭,再引起细胞膜出现新的电变化或其他细胞内功能变化,后者在Ca2+通道打开引起膜外Ca2+内流时甚为多见。
根据对Na+、K+、Ca2+三种离子的电压门控通道蛋白质进行的分子结构分析,发现它们一级结构中的氨基酸排列有相当大的同源性,说明它们属于同一蛋白质家族,与之有关的mRNA在进化上由同一个远祖基因演化而来。图2-8是与体内动作电位(见后)产生至关重要的Na+通道在膜内结构的模式图,它主要由一个较大的α-亚单位组成,分子量约260kd;有时还另有一个或两个小分子量的亚单位,分别称为β1和β2。但Na+通道的主要功能看来只靠α-亚单位即可完成。这个较长的α-单位肽链中包含了4个结构类似的结构域(domain,每个结构域大致相当于上述Ach门控通道中的一个亚单位,但结构域之间由肽链相连,是一个完整的肽链,应由一个mRNA编码和合成),而每个结构域中又各有6个由疏水性氨基酸组成的跨膜α-螺旋段(图示2-8,A);这4 个结构域及其所包含的疏水α-螺旋,在膜中包绕成一个通道样结构(图2-8,B)。现已证明,每个结构域中的第4个跨膜α-螺旋在氨基酸序列上有特点,即每隔两个疏水性氨基酸,就再现一个带正电荷的精氨酸或赖氨酸;这些α-螺旋由于自身的带电性质,在它们所在膜的跨膜电位有改变时会产生位移,因而被认为是该通道结构中感受外来信号的特异结构,由此再诱发通道“闸门”的开放;还有实验提示,每个结构域中的第2、第3个α-螺旋构成了该通道水相孔道的“内壁”;据测算,水相孔道内径最窄处横断面积约为0.3×0.5nm差不多刚能通过一个水化的Na+(图2-8,B)。
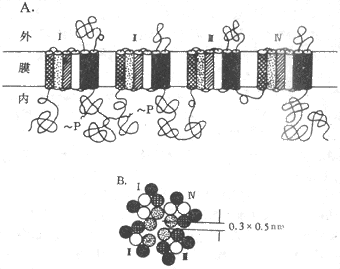
图2-8 电压门控Na+通道的分子结构示意图
A:构成电压门控Na+通道的α-亚单中的4个结构以及每个结构域中6个
α-螺旋在膜中存在形式平面 ~P表示磷酸化位点
B:4个结构域及其α-螺旋形成通道时的相对位置
(三)机械门控通道
体内存在不少能感受机械性刺激并引致细胞功能改变的细胞。如内耳毛细胞顶部的听毛在受到切和力的作用产生弯曲时,毛细胞会出现暂短的感受器电位,这也是一种跨膜信号转换,即外来机械性信号通过某种结构内的过程,引起细胞的跨膜电位变化。据精细观察,从听毛受力而致听毛根部所在膜的变形,到该处膜出现跨膜离子移动之间,只有极短的潜伏期,因而推测可能是膜的局部变形或牵引,直接激活了附近膜中的机械门控通道。
细胞间信道 还有一种通道,不是沟通胞浆和细胞外液的跨膜通道,而是允许相邻细胞之间直接进行胞浆内物质交换的通道,故称为细胞间通道。这种通道研究,是从缝隙连接超微结构观察开始的。在缝隙连接处相邻两细胞的膜仅隔开2.0nm左右,而且像是有某种物质结构把两者连接起来;将两侧细胞膜分离进行超微结构观察和分子生物学分析,发现每一侧的膜上都整齐地地排列着许多蛋白质颗粒,每个颗粒实际是由6个蛋白质亚单位(分子量各为25kd)构成的6聚体蛋白质,中间包绕一个水相孔道;构成颗粒的蛋白质和中心孔道贯穿所在膜的脂质双分子层;在两侧细胞膜靠紧形成细胞间的缝隙连接时,两侧膜上的各颗粒即通道样结构都两两对接起来,于是形成了一条条沟通两细胞胞浆的通路,而与细胞间液不相沟通。这种细胞间通道的孔洞大小,一般可允许分子量小于1.0~1.5kd或分子直径小于1.0nm的物质分子通过,这包括了电解质离子、氨基酸、葡萄糖和核苷酸等。这种缝隙连接或细胞间通道多见于肝细胞、心肌细胞、肠平滑肌细胞、晶状体细胞和一些神经细胞之间。缝隙连接不一定是细胞间的一种永久性结构;至少在体外培养的细胞之间的缝隙连接或其中包含颗粒的多少,可因不同环境因素而变化;似乎是细胞膜中经常有单方面装配好的通道颗粒存在,在两侧膜靠近并有其他调控因素存在时,就有可能实现对接,而在另一些因素存在时,两方面还可再分离。已对接的通道是否处于“开放”状态,也要受到多种因素的调控,例如当细胞内Ca2+、H+浓度增加时,可促使细胞间通道关闭。细胞间通道的存在,有利于功能相同而又密接的一组细胞之间进行离子、营养物质,甚至一些信息物质的沟通,造成它们进行同步性活动的可能性。
二、由膜的特异受体蛋白质、G-蛋白和膜的效应器酶组成的跨膜信号传递系统
这是另一类型的跨膜信号传递。最初是从对激素作用机制的研究开始的。60年代在研究肾上腺素引起肝细胞中糖原分解为葡萄糖的作用机制时,发现如果使肾上腺素单独和分离出的细胞膜碎片相互作用,可以生成一种分子量小、能耐热的物质,当把这种物质同肝细胞的胞浆单独作用时,也能引起其中糖原的分解,同肾上腺素作用于完整的肝细胞时有类似的效应。实验提示,在肾上腺素正常起作用时,它只是作用于肝细胞的膜表面。通过某种发生在膜结构中的过程,先在胞浆中生成一种小分子物质,后者再实现肾上腺素分解糖原的作用。这种小分子物质不久被证明是环-磷酸腺苷(即cAMP,环磷腺苷)。以后又陆续发现,很多其他激素类物质作用于相应的靶细胞时,都是先同膜表面的特异受体相结合,再引起膜内侧胞浆中cAMP含量的增加(有时是它的减少),实现激素对细胞内功能的影响。这样就把cAMP称作第二信使,这是相对于把激素分子这类外来化学信号看作第一信使而言的。
导致cAMP产生的膜结构内部的过程颇为复杂:它至少与膜中三类特殊的蛋白质有关。第一类是能与到达膜表面的外来化学信号作特异性结合的受体蛋白质,这是一些真正可以称作受体的物质。目前已用分子生物学的方法证明,它们是一些独立的蛋白质分子;已经确定的近100种这类受体,都具有类似的分子结构,也属于同一蛋白质家族:即它们都由约300~400个氨基酸残基组成,有一个较长的细胞外N-末端,接着在肽链中出现7个由22~28个主要为疏水性氨基酸组成的α-螺旋,说明这肽链至少要反复贯穿膜7次,形成一个球形蛋白质分子,还有一段位于膜内侧的肽链C-末端。目前认为,受体分子中第7个跨膜螺旋是能够识别、即能结合某种特定外来化学信号的部位;在受体因结合了特异化学信号而激活时,将进而作用于膜中另一类蛋白质,即G-蛋白质。
G-蛋白是鸟苷酸结合蛋白(guaninenucleotide-binding protein)的简称,也是存在于膜结构中的一类蛋白质家族,根据它们分子结构中少数氨基酸残基序列上的不同,已被区分出有数十种,但结构和功能极为相似。G-蛋白通常由α-、β-、和γ-3个亚单位组成;α-亚单位通常起催化亚单位的作用,当G-蛋白未被激活时,它结合了一分子的GDP(二磷酸鸟苷);当G-蛋白与激活了的受体蛋白在膜中相遇时,α-亚单位与GDP分离而又与一分子的GTP(三磷酸鸟苷)结合,这时α-亚单位同其他两个亚单位分离,并对膜结构中(位置靠近膜的内侧面)的第三类称为膜的效应器酶的蛋白质起作用,后者的激活(或被抑制)可以引致胞浆中第二信使物质的生成增加(或减少)。上述肾上腺素的作用,就是先由激素激活膜上相应的受体后,通过一种称为Gs(兴奋性G-蛋白)的G-蛋白的中介,激活了作为效应器酶的腺苷酸环化酶(图2-9箭头1),使胞浆中的ATP生成了起第二信使作用的cAMP(图2-9中箭头2)。由于第二信使物质的生成经过多级催化作用,少数几个膜外化学信号分子同受体的结合,就可能在胞浆中生成数目众多的第二信使分子,这是这种类型的跨膜信号传递的重要特点之一。
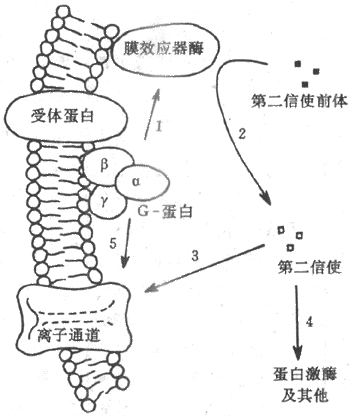
图2-9 由膜受体-G-蛋白-膜效应器酶组成的跨膜信号
传递系统和第二信使类物质的生成
目前发现膜的效应器酶并不只腺苷酸环化酶一种,因而第二信使物质也不只cAMP一种,如近年来还发现,有相当数量的外界刺激信号作用于受体后,可以通过一种称为Go的G-蛋白,再激活一种称为磷脂酶C的膜效应器酶,以膜结构中称为磷脂酰肌醇的磷脂分子为间接底物,生成两种分别称为三磷酸酰肌醇(IP3)和二酰甘油(DG)的第二信使,影响细胞内过程,完成跨膜信号传递。虽然如此,对应于细胞所能接受的多种刺激和与它们相对应的受体数目而言,膜内G-蛋白、效应器酶和最后生成的第二信使类物质的种类,还是相对地少得多。这说明,上述由膜中蛋白质酶促反应生成第二信使的途径,具有相当程度的“通用”性质。
由于上述这种跨膜信号传递的形式是在研究激素的作用机制时发现的,而且后来发现绝大多数肽类激素都是通过这一形式起作用的,因此曾一度错误地认为,这只是激素性化学信号跨膜信号传递方式。但近年的资料说明,事实并非如此:在神经递质类物质中,除了上述氨基酸类递质外,其余不论是小分子的经典递质还是后来发现的数量众多的神经肽类物质(目前已近50种),都主要是以在突触后细胞中产生第二信使类物质来完成跨膜信号传递的,这些第二信使物质通过在胞浆中的扩散,在膜的内侧面作用于某些特殊的离子通道(图2-9中箭头3),引起突触后膜较广泛而缓慢的电变化。最近证明,在视网膜信号转换过程中,光量子被作为受体的视色素如视紫红质(也具有7个跨膜α-螺旋的结构特点)吸收后,也是先激活称为Gt(转换蛋白)的G-蛋白,再激活作为效应器的磷酸二酯酶,使视杆细胞外段中cGMP的分解加强,最后使光刺激转变为外段膜的电变化(见第九章)。
上述两种主要的跨膜信号传递方式的作用过程,有以下几点值得注意。第一,这两种作用形式并不是绝对分离的,两者之间可以互相影响或在作用上有交叉。一些第二信使类物质可以调节某些电压门控通道和化学门控通道蛋白质的功能状态;而且被某种受体激活了的G-蛋白,有的不通过第二信使就能直接作用于膜结构中的通道结构(图2-9中的箭头5),如上述Gs激活时可以直接打开Ca2+通道。第二,对于许多外来化学信号分子,并不是一种化学信号只能作用于两种跨膜信号传递系统中的一种;以ACh为例,当它们作用于神经-肌接头处时,终板膜上有同它们作特异结合的化学门控通道;但当ACh作用于心肌或内脏平滑肌时,遇到的却是受体-G-蛋白-第二信使系统(受体称为M-型毒蕈硷型受体)。由此可见,同一种刺激信号通过何种跨膜信号传递系统起作用,关键因素在于靶细胞膜上具有何种感受结构;近年还发现,即便是M-型ACh受体,也可再区分出许多种亚型,有的亚型以cAMP为第二信使,有的以IP3和DG为第二信使。不同细胞甚或同一细胞的膜上具有对应于同一化学信号的不同受体型或其亚型,在跨膜信号传递中并不少见。近年来发现基本嗅觉刺激(大约是7种)全都是通过嗅上皮中不同的膜受体-第二信使系统起作用的,但在4种基本味觉刺激中,只有咸和酸刺激是通过细胞上相应的化学门控上通道起作用的,甜味物质是通过受体-第二信使系统起作用的,而苦味物质则因物质分子不同而分别通过通道和受体两种途径起作用。第三,跨膜信号传递的方式虽然相对地较少,但也不一定只限于上述两种。近年来有一些特殊的化学信号影响其靶细胞的方式受到广泛的重视,很可能成为跨膜信号传递的一种新类型;这就是发现胰岛素等一些肽类激素和其他与机体发育、生长、修复、增生、甚至细胞癌变有关的因子,如神经生长因子、表皮生长因子、血小板源生长因子、纤维母细胞生长因子、以及与细胞生成有关的集落刺激因子等,都是通过靶细胞表面一类称为酪氨酸激酶受体(tyrosine kinase riceptor)的蛋白质起作用的,这类受体结构简单,只有一个跨膜α-螺旋,当位于膜外侧的较长的肽链部分同特定的化学信号结合后,可以直接引起受体肽链的膜内段激活,使之具有磷酸激酶活性,通过使自身肽链和膜内蛋白质底物中的酪氨酸残基发生磷酸化,因而产生细胞内效应。这方面的新资料正在积累之中。
癌基因和跨膜信号传递近年发现与上述跨膜信号传递有关的一些蛋白质,如受体、G-蛋白、各种生长刺激因子和营养因子、以及各种蛋白激酶等,它们在细胞内的生物合成,是由人正常染色体中被称为细胞原癌基因(cellular proto-oncogene,进行表达时称细胞癌基因)的一类基因所编码和表达生成的。这些基因所以被称为原癌基因,是因为它们的硷基排列顺序同一些(近100种)能在动物引起肿瘤的病毒DNA(或称病毒癌基因,viral oncogene)的硷基排列顺序相一致。关于细胞癌基因与人类肿瘤发生的关系目前尚不清楚,但它们的正常表达产物,却是人体无时无刻不在进行着的各种跨膜信号传递过程所必需的。试设想,如果由于遗传和变异等原因使细胞不能合成结构和功能正常的G-蛋白,对人体将会有何等广泛而重要的影响!另外,在细胞原癌基因中,有一类可被胞浆中产生的第二信使等物质所激活,生成某种蛋白质;但它们在胞浆中生成后,一般又进入核内,进而诱导另一些基因进行表达。这类癌基因从激活到蛋白质生成,比一般基因表达为快,称为快速基因(或即早基因),而它们生成的蛋白质的作用则是激活另一些(可能在细胞功能活动中更重要)基因的表达,故快速基因的表达产物可称为转录调节因子或第三信使。所以称为第三信使是因为它们由第二信使类物质的作用而生成,而它们自身的作用又引起新的基因表达,生成一些可能对细胞结构和功能有较长远影响的蛋白质。这样外来信号(第一信使)不仅通过第二信使的合成在胞浆中引起一些即时反应,还可能通过第三信使引起细胞功能和结构长时间的适应性改变。
本文传播知识,如有侵权,请联系微信tunyi13166124885;本文非处方,如需治病,请联系医院。